Water is an essential yet scarce commodity for human life due to its many uses. Depending on their condition, bodies of water can improve or degrade living conditions. The Mvila watershed in southern Cameroon, which experienced archaic urbanization coupled with incivism and urban disorder, was the site of a study examining the relationship between the ecology of benthic macroinvertebrates and abiotic parameters. This study was conducted monthly from December 2018 to December 2019. It involved sampling benthic macroinvertebrates and collecting water samples using standard methods. For the most part, the physicochemical parameters remained within the standards prescribed by the water quality grid. Throughout the study, the Sounou stream had the highest abundance of benthic macroinvertebrates, followed by the Lo'o and Bengo streams. For the trophic groups, predators were the most abundant, while filter feeders were the least abundant. Canonical correspondence analysis revealed that certain physicochemical parameters had a negative influence on benthic macroinvertebrate taxa, while others had a positive impact on them. The parameters that negatively influenced them were temperature, pH, and conductivity, while the parameters that positively influenced them were suspended solids, colour, and turbidity. Finally, Shannon and Weaver's diversity indices and Pielou's evenness indicate a rich and diverse living environment with an equal distribution of organisms among the studied rivers. In conclusion, some environmental factors, such as temperature, pH, conductivity, and dissolved oxygen, influence the diversity and distribution of macroinvertebrates.
| Published in | Ecology and Evolutionary Biology (Volume 11, Issue 1) |
| DOI | 10.11648/j.eeb.20261101.11 |
| Page(s) | 1-11 |
| Creative Commons |
This is an Open Access article, distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution and reproduction in any medium or format, provided the original work is properly cited. |
| Copyright |
Copyright © The Author(s), 2026. Published by Science Publishing Group |
Ecology, Benthic Macroinvertebrates, Watershed, Taxa
Streams | Station code | Altitude | Geographic coordinates |
|---|---|---|---|
Metyi | M1 | 564 m | 02°52’22.7’’ N and 011°09’19.8’’ E |
M2 | 560 m | 02°52’50.6’’ N and 011°09’03.3’’ E | |
M3 | 559 m | 02°52’28.2’’ N and 011°09’22.0’’ E | |
Bengo | B1 | 578 m | 02°53’18.2’’ N and 011°09’28.2’’ E |
B2 | 577.5 m | 02°53’08.3’’ N and 011°09’24.9’’ E | |
B3 | 574 m | 02°53’03.9’’ N and 011°09’28.8’’ E | |
Sounou | S1 | 581 m | 02°52’36.7’’ N and 011°06’53.8’’ E |
S2 | 576 m | 02°52’41.8’’ N and 011°06’45.4’’ E | |
S3 | 570 m | 02°52’43.3’’ N and 011°06’41.4’’ E | |
Lo’o | L1 | 594 m | 02°53’48.7’’ N and 011°06’47.5’’ E |
L2 | 585 m | 02°53’53.7’’ N and 011°06’53.4’’ E | |
L3 | 579 m | 02°59’48.6’’ N and 011°06’58.7’’ E |
Metyi | Bengo | Sounou | Lo'o | Global | |
|---|---|---|---|---|---|
Temperature (°C) | 24.11±0.77 | 24.52±0.58 | 23.52±0.89 | 23.48±0.76 | 23.90±0.87 |
pH | 5.87±0.76 | 6.34±0.68 | 5.30±0.99 | 5.63±0.89 | 5.78±0.91 |
Oxygen (%) | 53.14±22.53 | 63.79±14.20 | 55.82±22.65 | 61.09±16.77 | 58.79±19.10 |
TDS (mg/L) | 12.18±5.71 | 63.85±14.44 | 11.28±4.09 | 12.56±4.42 | 24.97±23.99 |
Conductivity (µS/cm) | 25.38±9.06 | 124.36±26.24 | 25.38±6.82 | 27.69±8.42 | 50.71±45.15 |
Suspended Solid (mg/L) | 39.38±97.52 | 38.97±98.90 | 39.08±100.13 | 38.15±100.02 | 38.90±98.19 |
Turbidity (FTU) | 28.96±59.33 | 26.82±56.85 | 31.00±58.62 | 25.28±59.32 | 28.01±58.01 |
Orders | Metyi | Bengo | Sounou | Lo'o | ||||
|---|---|---|---|---|---|---|---|---|
N | N (%) | N | N (%) | N | N (%) | N | N (%) | |
Coleoptera | 618 | 47.1 | 543 | 32.8 | 328 | 10.3 | 361 | 21.6 |
Decapoda | 36 | 2.7 | 244 | 14.7 | 1811 | 56.9 | 512 | 30.6 |
Diptera | 16 | 1.2 | 6 | 0.4 | 30 | 0.9 | 21 | 1.3 |
Ephemeroptera | 41 | 3.1 | 54 | 3.3 | 299 | 9.4 | 179 | 10.7 |
Eulamellibranchia | 7 | 0.5 | 21 | 1.3 | 18 | 0.6 | 16 | 1.0 |
Heteroptera | 395 | 30.1 | 404 | 24.4 | 407 | 12.8 | 374 | 22.4 |
Caenogastropoda | 1 | 0.1 | 176 | 10.6 | 0 | 0.0 | 0 | 0.0 |
Odonata | 123 | 9.4 | 100 | 6.0 | 219 | 6.9 | 160 | 9.6 |
Plecoptera | 0.00 | 0.0 | 103 | 6.2 | 0 | 0.0 | 3 | 0.2 |
Trichoptera | 76 | 5.8 | 4 | 0.2 | 70 | 2.2 | 44 | 2.6 |
Oligochaeta | 0.00 | 0.0 | 0 | 0.0 | 3 | 0.1 | 3 | 0.2 |
Occurrence | Metyi | Bengo | Sounou | Lo'o | ||||
|---|---|---|---|---|---|---|---|---|
N | N (%) | N | N (%) | N | N (%) | N | N (%) | |
Rare taxa | 44 | 59.5 | 68 | 73.1 | 65 | 67.0 | 67 | 63.2 |
Accessory taxa | 16 | 21.6 | 12 | 12.9 | 17 | 17.5 | 23 | 21.7 |
Constant taxa | 8 | 10.8 | 6 | 6.5 | 5 | 5.2 | 6 | 5.7 |
Regular taxa | 5 | 6.8 | 5 | 5.4 | 5 | 5.2 | 6 | 5.7 |
Ubiquitous taxa | 1 | 1.4 | 2 | 2.2 | 5 | 5.2 | 4 | 3.8 |
Trophic groups | Metyi | Bengo | Sounou | Lo'o | Global | |||||
|---|---|---|---|---|---|---|---|---|---|---|
Absolute abundance | Number of families | Absolute abundance | Number of families | Absolute abundance | Number of families | Absolute abundance | Number of families | Absolute abundance | Number of families | |
Grazers / Shredders | 85 | 6 | 28 | 5 | 38 | 6 | 69 | 7 | 2651 | 12 |
Scrapers | 218 | 7 | 333 | 10 | 130 | 8 | 161 | 8 | 830 | 11 |
Detritivores | 68 | 8 | 247 | 10 | 1988 | 10 | 651 | 12 | 533 | 18 |
Filters | 81 | 2 | 24 | 2 | 82 | 3 | 59 | 3 | 246 | 4 |
Predators | 860 | 9 | 1022 | 13 | 944 | 14 | 732 | 14 | 3562 | 16 |
TDS | Total Dissolved Solids |
FTU | Formazing Turbidity Units |
µS/cm | microSiemens per Centimeter |
| [1] | Peter H., Gleick and Meena palaniapan (2010). Peak water-limits to freshwater withdrawal and use, proceding of the National Academy of Science: 11155 - 11162. |
| [2] | Oremo J., Orata F., Owino J. and Shivoga W. (2020). Assessment of available phosphates and nitrates levels in water and sediments of River Isiukhu, Kenya. Applied Ecology and Environmental Sciences 8: 119−127. |
| [3] | Onyando ZO., Lung’ayia H., Kigen CK., Shivoga WA. (2016). Dynamics in physico-chemical conditions along riparian land use gradients in River Isiukhu watershed, Western Kenya. International Journal of Environmental Biology 2: 53−60. |
| [4] | Lubanga HL. (2021). Spatial variability in water quality and macroinvertebrates assemblages across a disturbance gradient in the Mara River Basin, Kenya. Doctoral dissertation, University of Eldoret, Kenya. |
| [5] | Koskey JC., M’Erimba CM., Ogendi GM. (2021). Effects of land use on the riparian vegetation along the Njoro and Kamweti Rivers, Kenya. Open Journal of Ecology. 11: 807. |
| [6] | Mzungu E., Shivoga AW., Lukhabi KD. & Sifuna WA. (2025). Influence of physico-chemical parameters on benthic macroinvertebrate community dynamics in Isiukhu River, Kenya, African Journal of Aquatic Science, 1-14. |
| [7] | Nwaha M. (2023). Influence of certain environmental factors on the distribution of benthic macroinvertebrates in several rivers in the Mbila department in the South Region of Cameroon. Doctoral thesis in Animal Biology. University of Yaounde I, 181 pp. |
| [8] | Rosenberg D. M. and Resh V. H., 1993. Introduction to the freshwater biomonitoring and benthic macroinvertebrates. In: Rosenberg D. M. and Resh V. H. (eds.) freshwater biomonitoring and benthic macroinvertebrates, Chapman and all, New-York: 1-9. |
| [9] | Mbete V. C., Mbete P., Ibala-Zamba A. And Mamonekene V. (2021). Diversity of the benthic macroinvertebrate fauna in the Loya lagoon in southern Congo. Journal of Animal & Plant Sciences. 47(3): 8518-8526. |
| [10] | Foto Menbohan, S., Nwaha, M., Biram à Ngon, E. B., Dzavi, J., Boudem, R. C., Nangou, P. B. S. and Mbia, D. N. (2021) Water Quality and Benthic Macroinvertebrates of Tropical Forest Stream in South-West Region, Cameroon. International Journal of Progressive Sciences and Technologies, 25, 183-192. |
| [11] | Nwaha, M., Foto Menbohan, S., Nyame Mbia, D., Biram à Ngon, E., & Temgoua, Z. M. A. (2022). Water Quality Assessment of Some Forest Streams in the South Region of Cameroon. International Journal of Scientific Research and Engineering Development, 5(2): 1292-1302. |
| [12] | Dzavi, J., Menbohan, S. F., Mboye, B. R., Nwaha, M. and Biram à Ngon, E. B. (2022) Spatiotemporal Variation of Benthic Macroinvertebrates in Some Tropical Forest Streams of the Nyong Catchment (Cameroon). Open Journal of Applied Sciences, 12, 1210-1231. |
| [13] | Biram a Ngon, E. B., Ndjama, J., Chinche, S. B., Dzavi, J., Nwaha, M., Nyame Mbia, D. L., Epoundi, C. M., Betsi, W. C. & Menbohan S. F. (2024). Ecological Influence of Organic Pollution on the Distribution of Benthic Macroinvertebrates in Some Control Forest Watercourses in Cameroon. European Journal of Theoretical and Applied Sciences, 2(2), 1-5. |
| [14] | Min JK., Kong DS. (2020). Distribution patterns of benthic macroinvertebrate communities based on multispatial-scale environmental variables in the river systems of Republic of Korea. Journal of Freshwater Ecology 35: 323−347. |
| [15] | Mzungu E., Yakub S., Anyimba ES. (2022). Macroinvertebrates as bio-indicators of water quality in Omubira Stream, in Kakamega County, Kenya. International Journal of Fisheries and Aquatic Studies 10: 70−77. |
| [16] | Wanderi EW., Gettel GM., Singer GA., Masese FO. (2022). Drivers of water quality in Afromontane-savanna rivers. Frontiers. Environmental Science. 10: 972153. |
| [17] | Omoigberale MO., Ogbeibu AE. (2010). Environmental impacts of oil exploration and production on the macrobenthic invertebrate fauna of Osse River, Southern Nigeria. Research Journal Environmental Science 4: 101−114. |
| [18] | Andem AB., Ibor OR., Oku EE., Ekanem SB., Chukwuka AV., Adeogun AO. (2022). Urbanization gradients, riparian-loss and contaminant effects on macroinvertebrate distribution within a tropical river (Nigeria). Chemistry and Ecology 38: 503−526. |
| [19] | Raphahlelo ME., Addo-Bediako A., Luus-Powell WJ. (2022). Distribution and diversity of benthic macroinvertebrates in the Mohlapitsi River, South Africa. Journal of Freshwater Ecology 37: 145−160. |
| [20] | Mzungu E., Sifuna AW., Shivoga WA. (2023). Relationship between sediment grain sizes and macroinvertebrate distribution along the Isiukhu River, western Kenya. African Journal of Aquatic Science48: 274−286. |
| [21] | National Institute of Statistics (NIS) (2018). Statistical Yearbook of the Southern Region, 180p. |
| [22] | Ewane A. I. (2005). Structural analysis and pre-feasibility study of the Ebolowa Municipal Lake Development Project. Faculty of Sciences, University of Yaounde I, Cameroon. Specialised Higher Education Diploma Thesis. Environmental Sciences. 60 pp. |
| [23] | World Development Organisation (WDO) (2010): Regional Progress Report on the Millennium Development Goals for the Southern Region. |
| [24] | NCI. (1979). Topographic Map of Ebolowa and its Surroundings at a Scale of 1/50,000. Yaounde: National Cartographic Institute. Sheet 3D. |
| [25] | Rodier J., Legube B., Merlet N. et Coll. (2009). Water analysis: Natural waters, wastewater. Seawater. 9th edition. Dunod. Paris. 1526 p. |
| [26] | Stark J. D., Boothroyd K. G., Harding J. S., Maxted J. R. et Scarsbrook M. R. (2001). Protocols for Sampling Macroinvertebrates in Wadeable Streams. New Zealand Macroinvertebrates working group. Report no 1. 57 p. |
| [27] | Tachet H., Richoux P., Bournaud M. et UsseglioPolatera P. (2006). Systematic freshwater invertebrates. Biology and ecology. CNRS edition. Paris. 588 p. |
| [28] | Heidemann H. & Seidenbusch R. (2002). Larve et exuvies des libellules de France et d’Allemagne. 416p. |
| [29] | De Moor I. J., Day J. A. and De Moor F. C. (2003). Guides to the freshwater invertebrates of Southern Africa. Vol. 7: Insecta I. Ephemeroptera. Odonata & Plecoptera. Water Research Commission Report. No. TT 207/03. Pretoria. 288 p. |
| [30] | Stals R. & De Moor I. J. (2007). Guides to the Freshwater Invertebrates of Southern Africa. Volume 10: Coleoptera. Water Research Commission Report. No. TT 320/07. Pretoria. 263 p. |
| [31] | Dufrêne M. et Legendre P. (1997). Species assemblages and indicator species: the need for a flexible asymmetrical approach. Ecological Monographs, 67: 345-366. |
| [32] | Temgoua Zemo, M. A., Menbohan, S. F., Atchrimi, B. T., Betsi, W. C. N., Nwaha, M., Dzavi, J., Mavunda, C. A., Lactio, N. (2023). Effect of Anthropogenic Pressure on the Biodiversity of Benthic Macroinvertebrates in Some Urban Rivers (Yaoundé). Water. 15, 2383. |
| [33] | Mboye B. R. (2019). Diversity of benthic macroinvertebrates in the waterways of the Mabounie basin (Gabon): An essay on biotypology. Doctoral thesis. Faculty of Science, University of Yaoundé 1. 214 p. |
| [34] | Gwos Nhiomock S. R., Foto Menbohan S., Nyame Mbia D., Tchouapi Y. L., Biram A Ngon E. B. and Disso E. (2022). Biodiversity and water health status of four rivers in the East Cameroon region. GSC Biological and Pharmaceutical Sciences, 18(03), 226-241. |
| [35] | Ndourwe Far B., Edioh Pem F., Fosso Kengne L. P., Mogue Kamdem G., Betsi N. C. W., Gwos Nhiomock S. R., Foto Menbohan S. and Zebaze Togouet S. H. (2025). Biodiversity of Benthic Macroinvertebrates and water quality as a tool to the ecological study of three forest streams in the littoral zone (Cameroon). World Journal of Advanced Research and Reviews, 25(01), 1722-1734. |
| [36] | Eyre M. D., Foster G. N., Luff M. L. & Russhton S. P. (2006). The definition of British water beetle species pool (Coleoptera) and their relationship to altitude. Temperature, precipitation and land cover variables. Hydrobiologia. 560: 121-131. |
| [37] | Kabore I., Ouedraogo I., Tampo L., Oueda A., Moog O., Guenda W. et Melcher A. H. (2016 b). Composition and dynamic of benthic macroinvertebrates community in semi-arid area rivers of Burkina Faso (West Africa). International Journal of Biological and Chemical Sciences, 10: 1542-1561. |
| [38] | Kressou A. (2020). Diversity, structure of aquatic macroinvertebrate communities, and ecological integrity of aquatic environments in the Haut-Bandama Wildlife Reserve (north-central Ivory Coast). Doctoral thesis from Jean LOROUGNON GUEDE University (Ivory Coast) in Ecology, Biodiversity, and Evolution. 167p. |
| [39] | Prommi T. & Payakka A. (2015). Aquatic insect biodiversity and water quality parameters of streams in Northern Thailand. Sains Malaysiana, 44: 707-717. |
| [40] | Paredes-Agurto M. S. M., Ugaz Cherre A. F., Dioses J. M. M. and Garcia R. B. (2025). Aquatic Macroinvertebrate Diversity and Water Quality, La Gallega-Morropon Creek, Piura, Peru. Nature Environment and Pollution Technology. 23(4): 2397-2402. |
| [41] | Anderson N. H., Charles P. H. & Michael L. M. (1982). Effects of canopy, substrate composition and gradient on the structure of macroinvertebrate communities in Cascade Range stream of Oregon. Ecology, 63(3): 1840-1856. |
| [42] | Agadjihouede H., Chikou A. & Laleye P. (2009). Diversity and abundance of freshwater shrimp in the Grand-Popo lagoon (Lower Mono) in southern Benin. Proceedings of the 2nd UAC Colloquium on Science, Culture, and Technology, Agricultural Sciences: 462-469. |
| [43] | Toto Koume, Gouli Gore Bi, Claver Kemomadjehi Djirieoulou, Gervais Konan N’ZI and Dit Hervis Franck Gnonleba Boguhe (2018). Shrimp assemblage in the superior part of Cavally River (Coote d’Ivoire). International journal of innovation and applied study. 24(1): 379-388. |
| [44] | Tchakonte S., Ajeagah G. A., Diomande D., Camara A. I. et Ngassam P. (2014). Diversity, dynamic and ecology of freshwater snails related to environmental factors in urban and suburban streams in Douala-Cameroon (Central Africa), Aquat Ecol 48: 379-395. |
| [45] | Nyamsi Tchatcho N. L., Foto Menbohan S., Zebaze Togouet S. H., Onana Fils M., Adandedjan D., Tchakonte S., Yemele Tsago C., Koji E et Njine T. (2014). Yaounde Benthos Macroinvertebrate Multimetric Index (IMMY) for the biological assessment of water quality in rivers in the Central South Forest Region of Cameroon. European Journal of Scientific Research, 123: 412-430. |
| [46] | Aazami J., Esmaili-Sari A., Abdoli A., Sohrabi H. et Van den Brink P. J. (2015). Monitoring and assessment of water heath quality in Tajan River. Iran using physicochemical. Fish and macroinvertebrates indices. Journal of envirinnemnt heath Science and Enginneering. 13(2): 60-69. |
| [47] | Monoury E. (2013). Ecology of deposition zones in streams: contribution of invertebrates to litter decomposition and bioturbation processes. Doctoral thesis from the University of Toulouse 3 Paul Sabatier (France). 189 p. |
| [48] | Masese, F. O., Kitaka, N., Kipkemboi, J., Gettel, G. M., Irvine, K., & Mcclain, M. E. (2014). Macroinvertebrate functional feeding groups in Kenyan highland streams: evidence for a diverse shredder guild. Source: Freshwater Science, 33(2): 435-450. |
| [49] | Tenkiano N. (2017). Benthic macroinvertebrates and aquatic hyphomycetes: diversity and implications for ecosystem functioning in Guinea's waterways. Doctoral thesis from Toulouse 3 Paul Sabatier University (France). 226p. |
| [50] | Davies P. J., Wrigth I. A, Findlay S. J., Jonasson O. J. and Burgin S. (2010). Impact of Urban development on aquatic macroinvertebrate in south eastern Australia: degradation of in-stream habitats and comparison with non-urban steams. Aquatic Ecology, 44: 685-700. |
APA Style
Nwaha, M., Ngon, E. B. B. A., Dzavi, J., Mbongue, S. R., Epounde, A. C. M. (2026). Distribution of Benthic Macroinvertebrates in Some Rivers of the Ntem Basin in Southern Cameroon. Ecology and Evolutionary Biology, 11(1), 1-11. https://doi.org/10.11648/j.eeb.20261101.11
ACS Style
Nwaha, M.; Ngon, E. B. B. A.; Dzavi, J.; Mbongue, S. R.; Epounde, A. C. M. Distribution of Benthic Macroinvertebrates in Some Rivers of the Ntem Basin in Southern Cameroon. Ecol. Evol. Biol. 2026, 11(1), 1-11. doi: 10.11648/j.eeb.20261101.11
AMA Style
Nwaha M, Ngon EBBA, Dzavi J, Mbongue SR, Epounde ACM. Distribution of Benthic Macroinvertebrates in Some Rivers of the Ntem Basin in Southern Cameroon. Ecol Evol Biol. 2026;11(1):1-11. doi: 10.11648/j.eeb.20261101.11
@article{10.11648/j.eeb.20261101.11,
author = {Mathias Nwaha and Eric Belmond Biram a Ngon and Jean Dzavi and Serge Romeo Mbongue and Adele Carine Melle Epounde},
title = {Distribution of Benthic Macroinvertebrates in Some Rivers of the Ntem Basin in Southern Cameroon},
journal = {Ecology and Evolutionary Biology},
volume = {11},
number = {1},
pages = {1-11},
doi = {10.11648/j.eeb.20261101.11},
url = {https://doi.org/10.11648/j.eeb.20261101.11},
eprint = {https://article.sciencepublishinggroup.com/pdf/10.11648.j.eeb.20261101.11},
abstract = {Water is an essential yet scarce commodity for human life due to its many uses. Depending on their condition, bodies of water can improve or degrade living conditions. The Mvila watershed in southern Cameroon, which experienced archaic urbanization coupled with incivism and urban disorder, was the site of a study examining the relationship between the ecology of benthic macroinvertebrates and abiotic parameters. This study was conducted monthly from December 2018 to December 2019. It involved sampling benthic macroinvertebrates and collecting water samples using standard methods. For the most part, the physicochemical parameters remained within the standards prescribed by the water quality grid. Throughout the study, the Sounou stream had the highest abundance of benthic macroinvertebrates, followed by the Lo'o and Bengo streams. For the trophic groups, predators were the most abundant, while filter feeders were the least abundant. Canonical correspondence analysis revealed that certain physicochemical parameters had a negative influence on benthic macroinvertebrate taxa, while others had a positive impact on them. The parameters that negatively influenced them were temperature, pH, and conductivity, while the parameters that positively influenced them were suspended solids, colour, and turbidity. Finally, Shannon and Weaver's diversity indices and Pielou's evenness indicate a rich and diverse living environment with an equal distribution of organisms among the studied rivers. In conclusion, some environmental factors, such as temperature, pH, conductivity, and dissolved oxygen, influence the diversity and distribution of macroinvertebrates.},
year = {2026}
}
TY - JOUR T1 - Distribution of Benthic Macroinvertebrates in Some Rivers of the Ntem Basin in Southern Cameroon AU - Mathias Nwaha AU - Eric Belmond Biram a Ngon AU - Jean Dzavi AU - Serge Romeo Mbongue AU - Adele Carine Melle Epounde Y1 - 2026/02/11 PY - 2026 N1 - https://doi.org/10.11648/j.eeb.20261101.11 DO - 10.11648/j.eeb.20261101.11 T2 - Ecology and Evolutionary Biology JF - Ecology and Evolutionary Biology JO - Ecology and Evolutionary Biology SP - 1 EP - 11 PB - Science Publishing Group SN - 2575-3762 UR - https://doi.org/10.11648/j.eeb.20261101.11 AB - Water is an essential yet scarce commodity for human life due to its many uses. Depending on their condition, bodies of water can improve or degrade living conditions. The Mvila watershed in southern Cameroon, which experienced archaic urbanization coupled with incivism and urban disorder, was the site of a study examining the relationship between the ecology of benthic macroinvertebrates and abiotic parameters. This study was conducted monthly from December 2018 to December 2019. It involved sampling benthic macroinvertebrates and collecting water samples using standard methods. For the most part, the physicochemical parameters remained within the standards prescribed by the water quality grid. Throughout the study, the Sounou stream had the highest abundance of benthic macroinvertebrates, followed by the Lo'o and Bengo streams. For the trophic groups, predators were the most abundant, while filter feeders were the least abundant. Canonical correspondence analysis revealed that certain physicochemical parameters had a negative influence on benthic macroinvertebrate taxa, while others had a positive impact on them. The parameters that negatively influenced them were temperature, pH, and conductivity, while the parameters that positively influenced them were suspended solids, colour, and turbidity. Finally, Shannon and Weaver's diversity indices and Pielou's evenness indicate a rich and diverse living environment with an equal distribution of organisms among the studied rivers. In conclusion, some environmental factors, such as temperature, pH, conductivity, and dissolved oxygen, influence the diversity and distribution of macroinvertebrates. VL - 11 IS - 1 ER -
Department of Biomedical Sciences, Univeristy of Bertoua, Bertoua, Cameroon
Research Centre for Water and Climate Change, Institute of Geological and Mining Research, Yaounde, Cameroon
Research Centre for Water and Climate Change, Institute of Geological and Mining Research, Yaounde, Cameroon;Department of Animal Biology and Physiology, University of Yaounde I, Yaounde, Cameroon
Department of Biomedical Sciences, Univeristy of Bertoua, Bertoua, Cameroon
Department of Animal Biology and Physiology, University of Yaounde I, Yaounde, Cameroon
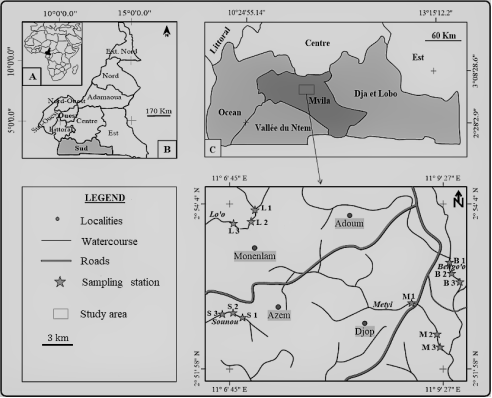
Figure 1. Sampling station in different streams in Ntem watershed [24].
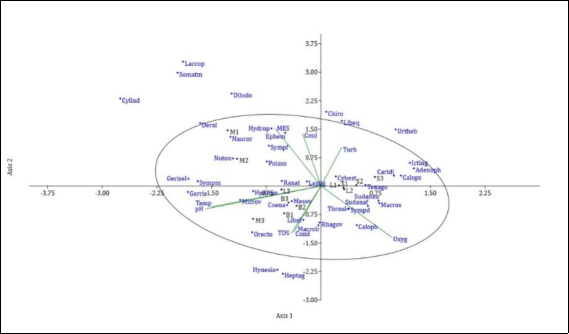
Figure 2. Canonical Correspondence Analysis (CCA) of ubiquitous, regular, and constant macroinvertebrate taxa in relation to abiotic parameters. Macrob = Macrobrachium; Caridi = Caridina; Ranat = Ranatra; Poisso = Poissonia; Hydrom = Hydrometra; Cylind = Cylindrostethus; Tenago = Tenagogonus; Rhagov = Rhagovelia; Mesov = Mesovelia; Microv = Microvelia; Naucor = Naucoris; Deral = Derallus; Orecto = Orectogyrus; Laccop = Laccophilus; Heptag = Heptagenia; Ephem = Ephemerella; Thraul = Thraulus; Hydrop = Hydropsyche; Lestig = Lestinogomphus; Ortheb = Orthetrum; Sympd = Sympetrum; Coena = Coenagrion; Temp = Temperature; pH = Hydrogen potential; Oxyg = Dissolved oxygen; TDS = Total Dissolved Solids; Cond = Electrical conductivity; MES = Suspended solids; Turb = Turbidity.
Information

